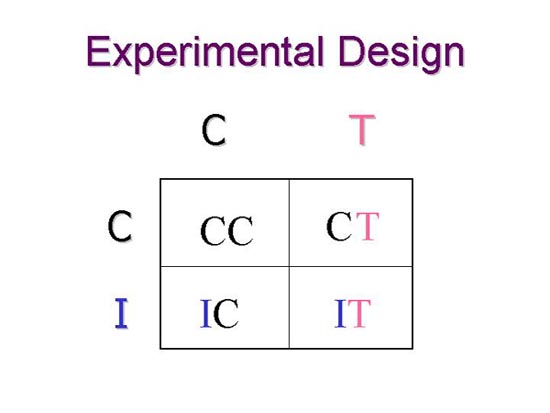
We used a two-by-two design with testosterone level (T) and immunochallenge (I) as the two treatment variables. Corresponding controls (C) were established. The result was four groups of adult male white-footed mice.
Funded by:
Howard Hughes Medical
Institute
Committee
on Institutional Studies and Research, MSU
According to the
"winter immunoenhancement hypothesis," some species of mammals exhibit
an
increase in immune function prior to winter, resulting in improved
defense from pathogens during harsh winter conditions. It has
also been suggested that the enhancement of immune function is
possible,
in part, because of a simultaneous reduction in reproductive
activity.
As a result, energy and nutrients can be re-allocated to support immune
functions. The occurrence of energy re-allocation would indicate
that trade-offs exist between energy use for immune and reproductive
functions.
We demonstrated previously that a mild immune challenge was associated
with a decrease in testes mass in white-footed mice. To further
investigate the ability of animals to accommodate the energetic costs
of immune and reproductive functions, we examined the effects of
testosterone
and immunochallenges on energy budgets, again using white-footed
mice.
Understanding of the trade-offs that may exist between these two
systems
is important to understanding the potential impacts of the increasing
incidence of infectious diseases and parasites in wild animal and human
populations worldwide.
|
We used a two-by-two design with testosterone level (T) and immunochallenge (I) as the two treatment variables. Corresponding controls (C) were established. The result was four groups of adult male white-footed mice. |
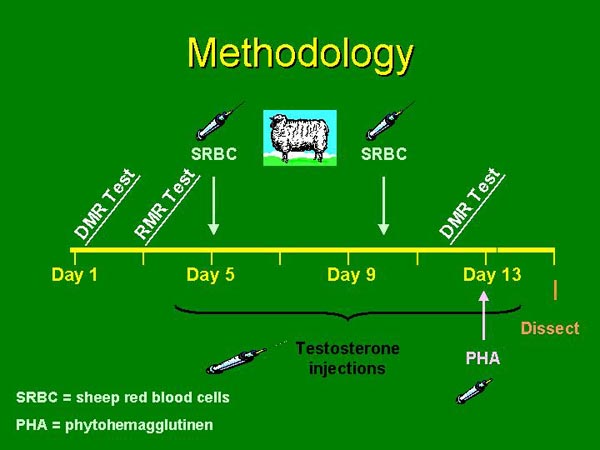
| To determine patterns of energy allocation in each group of
mice, we measured resting and daily metabolic rates before and after
injections of sheep red blood cells (SRBC) and phytohemagglutinen
(PHA).
SRBC challenged the humoral component of the immune system and PHA
challenged
the cell-mediated
component. Control animals were injected with saline. The
testosterone-treated animals were injected with testosterone propionate
daily and the controls
were injected with saline. At the end of the experiment the
animals were dissected. |
|
|
 |
 |
 |
 |
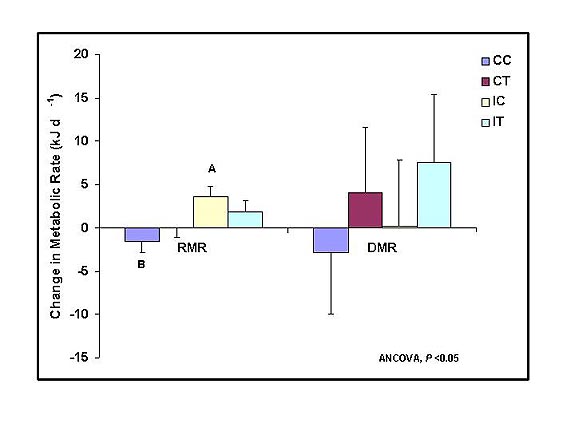
| Resting metabolic rate (RMR) was
significantly greater by the end of the experiment in animals that were
immunochallenged (IC) compared with the control group (CC).
Elevated testosterone was associated with suppression of the
increase in resting metabolic rate (IT). Daily metabolic rate
(DMR) did not
increase significantly during the immunochallenges. |
|
 |
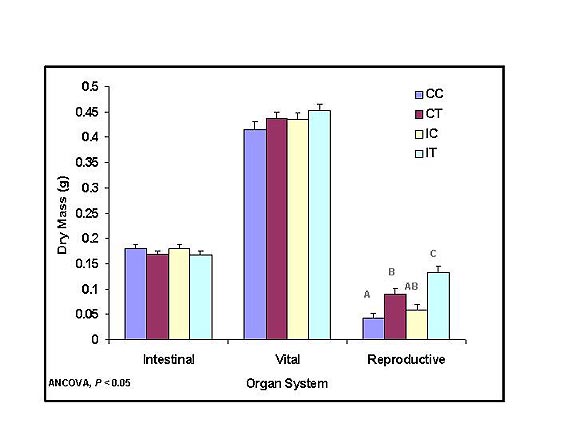
Surprisingly, the reproductive organs (seminal vesicles and testes) were significantly larger in males that were immunochallenged (IC, IT) compared with the control males (CC, CT). These data indicated two important results. First, the animals were capable of mounting an immune response and increasing energy allocation to reproductive organs. Second, the increase in resting metabolic rate (graph above) was attributable to the increase in immune function rather than the increase in reproductive organ masses. |
|
 |
CONCLUSIONS:
|